

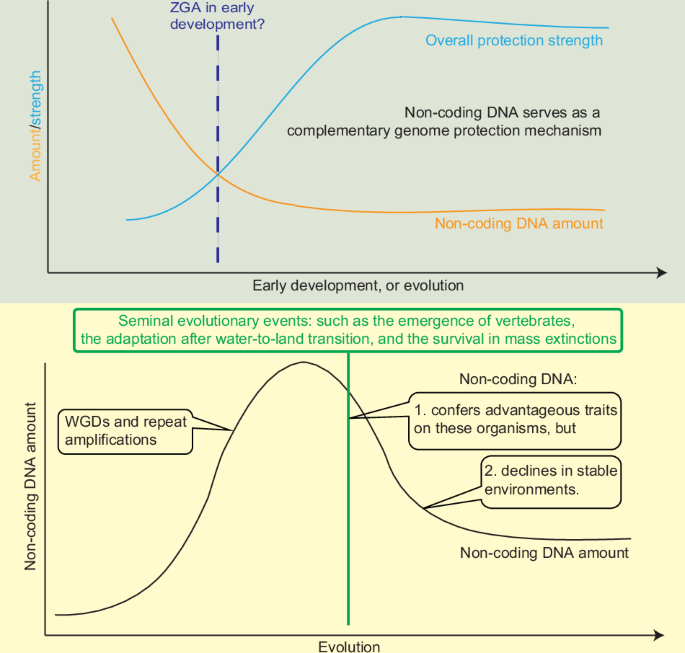
It is generally accepted that non-coding DNA constitutes the vast majority of most eukaryotic genomes and is concentrated at the nuclear periphery and nucleolar surface. Building upon this spatial organization, we have previously proposed that this layer of abundant, peripherally localized non-coding DNA functions as a 3D buffer that transiently absorbs or permanently excludes DNA damage to protect the genome and the relatively central exome from external and internal mutagens in somatic cells. This review explores the potential role of non-coding DNA as a physical barrier in genome safeguarding during early developmental stages and major evolutionary transitions. During gametogenesis and early embryogenesis the barrier is first provided by abundant non-coding DNA; as heterochromatin matures from non-coding DNA, many species programmatically eliminate the now-redundant non-coding DNA. Across evolution, whole-genome duplications and repeat amplification expand the shield, facilitating major evolutionary transitions such as vertebrate origins, water-to-land colonization and survival through mass-extinction crises. Conversely, in stable ecosystems selection favors genome streamlining: redundant non-coding DNA is lost to reduce fitness costs. Thus, the dosage of non-coding DNA is negatively correlated with the strength of apomorphic safeguards (adaptive immunity, viviparity) and positively correlated with ecological or developmental stress. By integrating comparative genomics, 3D nuclear architecture and evolution, we unify a single conceptual framework: non-coding DNA acts as a malleable fortress whose thickness is tuned to the variable need for genome protection during both ontogeny and phylogeny. This perspective offers new explanatory power for the accumulation or loss of non-coding DNA and can predict genome-size trajectories. Aburomia R, Khaner O, Sidow A (2003) Functional evolution in the ancestral lineage of vertebrates or when genomic complexity was wagging its morphological tail. J Struct Funct Genom 3(1-4):45–52. Aguirre-Lavin T, Adenot P, Bonnet-Garnier A, Lehmann G... [27352 chars]
ADVERTISEMENT

ADVERTISEMENT
2026 Copyright © News-Matic.com. All Rights Reserved



